








Plant cell apoptosis
 source:ELK Biotechnology
source:ELK Biotechnology date:2022-05-19
date:2022-05-19 views:2515
views:2515
Cell death can be driven by a genetically programmed signalling pathway known as programmed cell death (PCD). In plants, PCD occurs during development as well as in response to environmental and biotic stimuli. Our understanding of PCD regulation in plants has advanced significantly over the past two decades; however, the molecular machinery responsible for driving the system remains elusive. Thus, whether conserved PCD regulatory mechanisms include plant apoptosis remains enigmatic. Animal apoptotic regulators, including Bcl-2 family members, have not been identified in plants but expression of such regulators can trigger or suppress plant PCD. Moreover, plants exhibit nearly all of the biochemical and morphological features of apoptosis. One difference between plant and animal PCD is the absence of phagocytosis in plants. Evidence is emerging that the vacuole may be key to removal of unwanted plant cells, and may carry out functions that are analogous to animal phagocytosis.Multicellular organisms have intrinsic and extrinsic programs for cell suicide. Programmed cell death (PCD) is regulated by death receptors or intrinsic genes, and these signals direct cells to resist pathogens or environmental damage by removing themselves during development. PCD has been found to be characteristic of prokaryotes such as bacteria and yeast, the existence of chloroplasts and mitochondria in prokaryotic cells is related to PCD in plants, while they are the main sources of reactive oxygen species, and animal organelles play a crucial role in PCD.
Definition of cell death
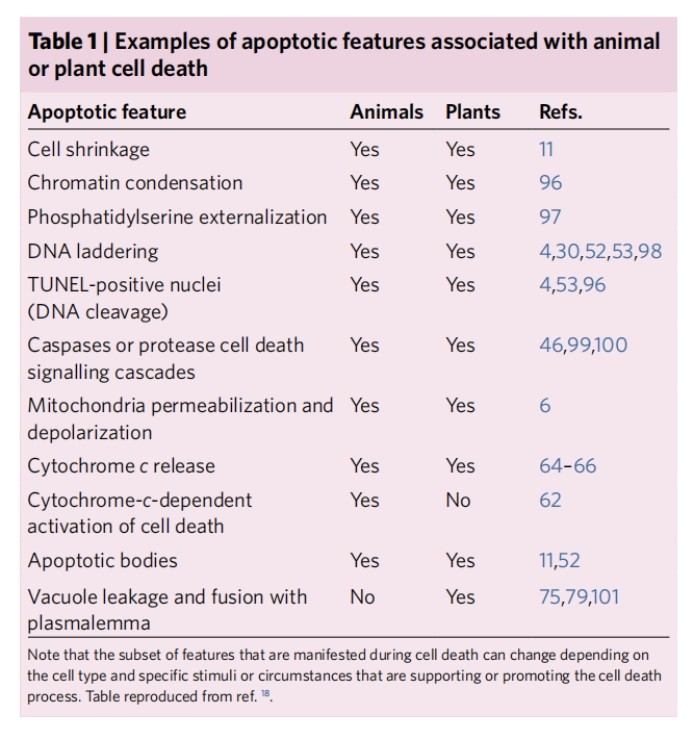
There are many ways cells die, but not all cell death regulators function the same. PCD is a continuous process of cell death.
Apoptosis is one of the most intensively studied forms of PCD and in animals it has two main pathways, the external pathway and the intrinsic pathway, distinguished by the source of the death signal. The extrinsic pathway begins with cell surface receptors that bind to death ligands and trigger intracellular signalling pathways that lead to death. In contrast, the intrinsic pathway is usually controlled by signals emanating from the mitochondria. Cells undergoing apoptosis exhibit typical morphological and biochemical features including chromatin condensation, DNA cleavage (ladder), TUNEL (terminal deoxynucleotidyl transferase dUTP end-labelling) positive nuclei, apoptotic vesicle formation, phosphatidylserine externalisation and macrophage phagocytosis, and from these initial hallmark responses, apoptosis appears to trigger in a linear fashion an ordered disassembly of the cell and eventual death (Figure 1). It is clear that there are multiple ways in which cells can be programmed to die, including non-apoptotic forms of PCD such as iron death, cell scorching and necrosis. Thus, PCD can be considered as a continuum, a fluid target that may increase in number and complexity.
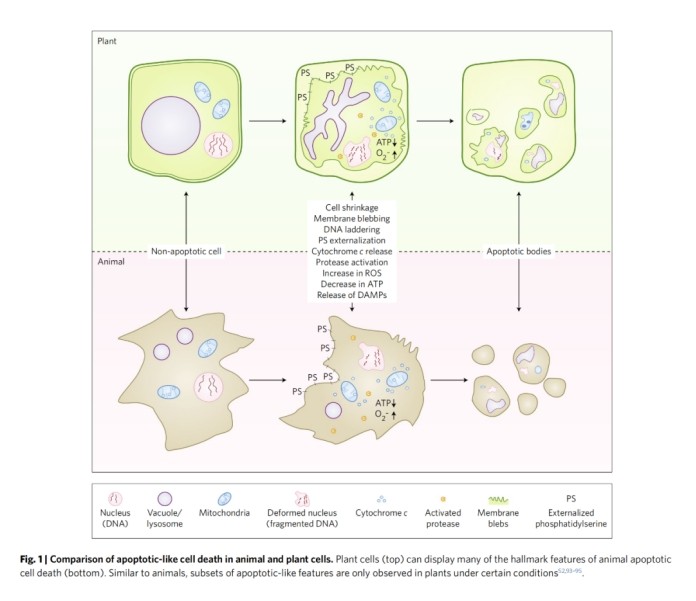
(Figure 1)
Plant PCDs and animal PCDs are similar but different
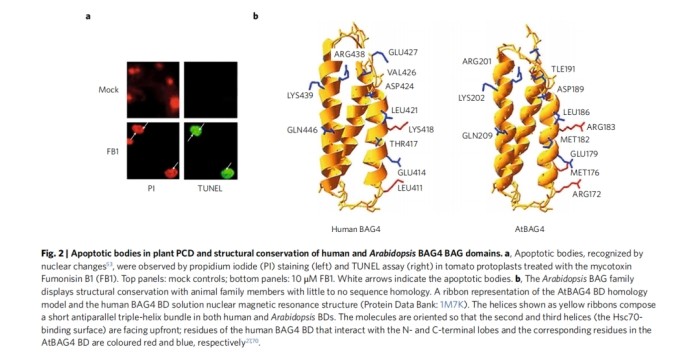
Apoptosis in plants
The production of apoptotic bodies is unique to animal cell apoptosis. However, apoptotic-like bodies have been observed in plant cells subjected to both abiotic and biotic-induced stimuli. Under some special temperature stimuli, some plants Cells also show apoptosis-like body formation. Plant genomes may also possess genes encoding functional equivalents of anti-apoptotic Bcl-2 family proteins with little or no sequence homology to their corresponding proteins in animal cells. Similar only in structure, having the same functional domains of the molecule. Mouse Bax expressed in plants is constitutively localized to mitochondria while leading to the release of cytochrome c.
The role of plant vacuoles in clearing cellular debris
In the absence of a phagocytic system, plant vacuoles play a key role in the removal of unneeded plant cells. Phagocytosis of cytoplasmic material by the vacuole is associated with vesicular cell death (VCD), which leads not only to the phagocytosis of protoplasts but also to the degradation of plant cell walls.
Vesicle rupture in which hydrolytic enzymes are expelled into the cytoplasm contains papain-like cysteine proteases (PLCPs), RD21 being one of the main proteases responsible for regulating oxalic acid (OA)-induced apoptotic-like cell death activity, and excess RD21 accelerates apoptosis.
PCD regulators in animals act in plants
The core regulator gene of animal PCD is at a low level in the plant genome. By studying the effects of ectopic expression of animal pro- and anti-apoptotic proteins in transgenic plants, Ced-9, an animal Bcl-2 homolog, is localised to the inner mitochondrial membrane. The expression of ced-9 in plant cells affects nodule formation and reduces apoptosis by regulating the vesicle trafficking pathway and interfering with autophagy.
Another group of PCD regulators with limited sequence homology in plants is the inhibitors of apoptosis proteins (IAPs). Proteins contain the BIR domain, most of which also contain the c-terminal RING domain, a structurally distinct zinc finger domain that attenuates caspase activity. No BIR domain protein has been found in plants. On the contrary, plants encode IAP-like proteins (ILPs) that lack the BIR domain but contain the C-terminal RING domain.The RING domain has E3 ligase activity, which is the core region of proteasomal regulatory activity. Arabidopsis IAP-like proteins (AtILPs) can also slow down apoptosis by inhibiting caspase-3 activity.
Conclusion and Development
Although great progress has been made in recent years in characterizing plant PCD; however, the mechanism of plant cell death remains unclear. Why animal oncogenes confer disease resistance in plants/fungi.
(1) Presence of apoptotic-like bodies.
(2) The importance of structural similarity, which maintains functional consistency.
Future research on apoptosis-like proteins in plants is continuing, and there will be more use of the term "apoptosis-like" in plant research.
Literature source:
Reassessing apoptosis in plants | Nature Plants

 RETURN
RETURN